
Evolución paralela del código genético en genomas mitocondriales de artrópodos
Una de las propiedades más notables del código genético es que es el mismo en la mayoría de los organismos. Esta notable conservación sugiere que fue establecido tempranamente en la evolución de la vida en la Tierra, antes de la división de los tres principales dominios [1] y ha permanecido constante desde entonces. Este código no es aleatorio, varios estudios han relacionado la forma del código genético canónico con propiedades estereoquímicas de aminoácidos y codones, minimización del impacto de mutaciones y relaciones biosintéticas entre diferentes aminoácidos

El código genético provee la tabla de traducción necesaria para transformar la información contenida en el ADN al lenguaje de proteínas. En esta tabla, la correspondencia entre cada codón y cada aminoácido es establecida: el ARNt es el adaptador principal que une a ambos. Si bien el código genético es casi universal, diversas variantes de este código han sido descriptas en un amplio rango de sistemas nucleares y de organelas, especialmente en la mitocondria de animales pluricelulares.
Estas variantes son generalmente encontradas buscando posiciones conservadas que con frecuencia codifican para un aminoácido alternativo en nuevas especies. Hemos ideado un preciso método computacional para automatizar estas comparaciones, y lo hemos probado con el genoma mitocondrial de 626 animales pluricelulares. Nuestros resultados indican que varios artrópodos tienen un nuevo código genético y traducen el codón AGG como lisina en vez de serina (como en el código genético mitocondrial de los invertebrados) o arginina (como en el código genético estándar). Hemos investigado la evolución del código genético en artrópodos y encontramos varios hechos de evolución paralela en los cuales el codón AGG era reasignado entre serina y lisina. Nuestro análisis también reveló una correlación entre los códigos genéticos de artrópodos y el ARNt-Lis/Ser, lo cual muestra puntos de mutaciones específicos en los anticodones. Estas mutaciones bastante simples, junto con el bajo uso del codón AGG, pueden explicar la repitencia de las reasignaciones AGG.
Precisión de la predicción computacional del código genético
Para probar la validez de nuestro enfoque, comparamos las asignaciones automáticas con los códigos genéticos provistos en GenBank para cada especie. Asumiendo que las predicciones que no son concordantes con GenBank son erróneas (una suposición que es usual, pero no siempre correcta) fuimos capaces de estimar la precisión del método.
En la Figura 1 graficamos la cantidad de predicciones concordantes/no concordantes así como el número de codones no predichos (aquellos que no fueron observados en las posiciones debajo del umbral de entropía) para cuatro umbrales de entropía diferentes (S).

Un nuevo código genético en artrópodos
Extraordinariamente, algunas asignaciones no concordantes ocurrieron con un número intermedio de observaciones de codones y una alta frecuencia de aminoácidos (algunos de los triángulos amarillos en la figura 2), en una región en la cual las asignaciones fueron casi todas correctas. Todas estas observaciones corresponden a cambios en el codón AGG, el cual se traduce a Ser de acuerdo al Código Genético Mitocondrial de los Invertebrados (IMGC), pero fue predicho que se transcribía como Lys en la región de alta confianza de varios artrópodos.
En otros artrópodos se predijo el mismo cambio, pero localizado en una región de la Figura 2 en la cual las asignaciones correctas y las incorrectas son frecuentes (los restantes triángulos amarillos). Esta asignación particular de codones era consistente si usábamos para el análisis a todos los metazoarios (n = 626) o solo artrópodos (n = 92), eliminando la posibilidad de que las asignaciones de artrópodos fueran pobremente predichas debido a la distancia del resto de los metazoarios.
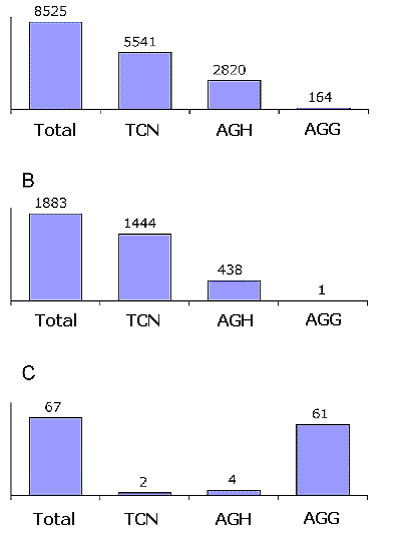
Estas asignaciones de codones fueron también consistentes bajo diferentes umbrales de conservación (datos no publicados). La predicción de que AGG codifica Lys fue también la única asignación no concordante que se repitió en varias especies. Estas observaciones sugieren fuertemente que AGG a Lys, es una nueva reasignación de codón, previamente no observada. De acuerdo a nuestros resultados del conjunto de datos de artrópodos, 24 especies traducen AGG a Lys y 34 especies lo hacen a Ser. Para 18 especies el significado de AGG no puede ser predicho, y 16 especies no utilizan AGG para nada. En las especies donde se predijo que AGG se traduce en Lys, los codones TCN y AGH (AGC, AGT, AGA) están claramente asociados con las columnas alineadas donde Ser está conservada en un porcentaje mayor al 80%, mientras que el codón AGG es claramente asociado con las columnas conservadas de Lys (Figura 3).
Este último efecto es ejemplificado por el crustáceo Speleonectes tulumensis. Esta especie tiene un total de 17 codones AGG, de los cuales nueve aparecen en posiciones por debajo del umbral de entropía. Ocho de estos nueve codones aparecen en posiciones donde la Lys está conservada en más del 80%. La probabilidad de que este suceso sea una causalidad es muy pequeña: hay 37 columnas alineadas de un total de 2443 con más del 80% de Lys, y la probabilidad de que después se coloquen aleatoriamente nueve codones AGG, al menos ocho de ellos ocupando estas columnas de Lys es:
P(X =8) = 1,5 x 10-17
Evolución del código genético en artrópodos
Para entender más profundamente el origen y la distribución del nuevo código genético; examinamos su evolución a través de la filogenia de los artrópodos. Sin embargo, la filogenia de los principales linajes de artrópodos es polémica. Nosotros utilizamos un árbol filogenético de consenso que ensamblamos a partir de diferentes fuentes [19-26] para reflejar mejor el conocimiento actual de las relaciones entre artrópodos (figura 4). Las politomías fueron introducidas en varios casos donde existían incertezas: la posición relativa de los hexápodos con respecto a los miriápodos y crustáceos (las hipótesis Atelocerata y Pancrustacean respectivamente) [22], la mono/parafilia de los hexápodos (dependiendo de la posición relativa de Ellipura con respecto a los insectos) [20,21,27], la monofilia de crustáceos [24], y las relaciones entre diferentes clases de crustáceos. La filogenia de las diferentes ordenes de insectos estuvo basada principalmente en referencias [19].
La reconstrucción más moderada [28] de la evolución del código genético en este árbol indica claramente que el código genético mitocondrial de los artrópodos cambio en muchas ocasiones (Figura 4). Inesperadamente, los cambios ocurren dentro de los grupos mayoritarios y minoritarios de artrópodos. Por ejemplo, en la orden Hemiptera, Euhemiptera (Philaenus spumarius, Triatoma dimidiata) lee AGG como Lys, pero Sternorrhyncha (Aleurochiton aceris, Bemisia tabaci, Tetraleurodes acaciae, Trialeurodes vaporariorum, Neomaskellia andropogonis, Aleurodicus dugesii, Schizaphis graminum) lee AGG como Ser. Un patrón similar fue observado en la subclase de chelicerate, Acari. Interesantemente, y sin tener en cuenta la inclusión de un grupo de especies con el IMGC, la reconstrucción del estado ancestral indica que el código genético que traduce AGG como Lys es el más antiguo. Confirmamos que la reconstrucción del estado ancestral da el mismo resultado cuando resolvemos las politomías de diferentes maneras (de acuerdo filogenias de artrópodos recientemente publicadas [20–22,24,29]), y cuando enraizamos el árbol con un grupo que tiene el IMGC.
En todas, excepto una, de las combinaciones alternativas que analizamos se predijo que claramente que el estado ancestral traducía AGG como Lys.
En ningún caso se predijo que el IMGC sea el código ancestral de los atrópodos. Por lo tanto decidimos nombrar este novedoso código genético como Ancestral Arthropod Mitochondrial Genetic Code (AAMGC) para diferenciarlo del IMGC en el cual AGG es traducido como Ser. La naturaleza ancestral de este AAMGC está fuertemente sostenida por la observación de que es encontrado en varios linajes de artrópodos, que se consideran divergencias tempranas. Por ejemplo, en el linaje basal más ampliamente aceptado de Chelicerata (cangrejo herradura -Merostomata-) [23], Hexapoda (Ellipura) [19], e Insecta (Thysanura) [19], todos se predijeron utilizando el AAMGC.
Este es también el caso de las clases Remipedia y Branchiopoda, que han sido a menudo propuestos para estar entre los crustáceos más primitivos [30]. Los Miriápodos también usan el AAMGC.
Referencia











No hay comentarios.: